
Anti-Ovarian Cancer Conotoxins Identified from Conus Venom
by
 , and
, and
Shuang Ju
1,
Yu Zhang
1,
Xijun Guo
1,
Qinghui Yan
1,
Siyi Liu
1,
Bokai Ma
2,
Mei Zhang
2,
Jiaolin Bao
1,
Sulan Luo
1,3 and
Ying Fu
1,* 1
Key Laboratory of Tropical Biological Resources of Ministry of Education, School of Pharmaceutical Sciences, Hainan University, Haikou 570228, China
2
Beijing Key Laboratory of Organic Materials Testing Technology & Quality Evaluation, Institute of Analysis and Testing, Beijing Academy of Science and Technology, Beijing 100094, China
3
Medical School, Guangxi University, Nanning 530004, China
*
Author to whom correspondence should be addressed.
Molecules 2022, 27(19), 6609; https://doi.org/10.3390/molecules27196609
Submission received: 29 August 2022
/
Revised: 16 September 2022
/
Accepted: 30 September 2022
/
Published: 5 October 2022
Abstract
:Conotoxins constitute a treasury of drug resources and have attracted widespread attention. In order to explore biological candidates from the marine cone snail, we isolated and identified three novel conopeptides named as Vi14b, Vi002, Vi003, three conotoxin variants named as Mr3d.1, Mr3e.1, Tx3a.1, and three known conotoxins (Vi15a, Mr3.8 and TCP) from crude venoms of Conus virgo, Conus marmoreus and Conus texile. Mr3.8 (I-V, II-VI, III-IV) and Tx3a.1 (I-III, II-VI, IV-V) both showed a novel pattern of disulfide connectivity, different from that previously established for the µ- and ψ-conotoxins. Concerning the effect on voltage-gated sodium channels, Mr3e.1, Mr3.8, Tx3a.1, TCP inhibited Nav1.4 or Nav1.8 by 21.51~24.32% of currents at semi-activated state (TP2) at 10 μmol/L. Certain anti-ovarian cancer effects on ID-8 cells were exhibited by Tx3a.1, Mr3e.1 and Vi14b with IC50 values of 24.29 µM, 54.97 µM and 111.6 µM, respectively. This work highlights the role of conotoxin libraries in subsequent drug discovery for ovarian cancer treatment.
1. Introduction
When comparing with other types of cancer, ovarian cancer is the eleventh most common in women, the fifth-most common cause of cancer-related death in women, and the most fatal gynecologic malignant cancer for females [1,2,3]. Hidden deep and often known as a silent killer, ovarian cancer is frequently not diagnosed (over 70%) until the disease has advanced to the late stage, since its symptoms are vague and difficult to detect [4,5]. In the year 2022 alone, 19,880 new cases and 12,810 deaths related to ovarian cancer were estimated by the National Cancer Institute of the United States [3]. Treatment of ovarian cancer is generally performed by a combination of debulking surgery and subsequent chemotherapy with the increasing use of immunotherapy and targeted therapies such as bevacizumab and PARP (poly ADP-ribose polymerase) inhibitors [6,7,8]. Paclitaxel/carboplatin and doxorubicin/carboplatin regimens are often used as first-line chemotherapeutic agents [2]. However, failure of chemotherapy may occur due to drug resistance problems and toxicity [9]. Herein, alternative anti-ovarian cancer drug candidates are urgently needed for the treatment of ovarian cancer.
Conotoxins (Ctx) are a class of active polypeptides secreted by cone snails (a carnivorous mollusk living in tropical oceans) and are mainly used for prey capture and defense against natural enemies [10,11,12]. Conotoxins usually consist of 10–40 amino acids and contain 0–8 cysteine (Cys) residues, which can form 0–4 pairs of intramolecular disulfide bridges. Conotoxins can specifically target on diverse crucial receptors (ligand-gated ion channels, voltage-gated ion channels, G protein-coupled receptors, and neurotransmitter transporters, etc. [13,14,15]) related to tough diseases, such as neuralgia, addiction, epilepsy, mental illness, cancer and other intractable diseases [16,17,18,19]. To date, more than seven contoxins, including ω-MVIIA (chronic pain, marketed), ω-CVID (analgesia, phase IIa), contulakin-G (analgesia, phase I), conantokin-G (analgesia/anti-epileptic, phase Ib), χ-MrIA (analgesia, phase IIa), α-Vc1.1 (analgesia, phase II, terminated), μO-MrVIB (analgesia, phase II), etc., have advanced to the clinical research stage [20,21]. Conoxin ω-MVIIA, which targets N-type human Cav2.2 channels, has been developed as an FDA-approval analgesic drug Ziconotide (Prialt) for severe chronic pain since its launch in 2004 [22]. Cone snail venom is a huge drug resource, since it is estimated that cone snails can generate up to one million different bioactive peptides [23]. However, less than 0.1% of these useful peptides have been structurally and functionally characterized [15,18,21]. Herein, conotoxin has attracted extensive attention and becomes a hot spot in new drug development.
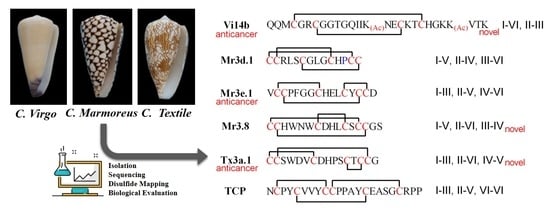
In order to explore biological candidates from marine cone snails, we isolated and identified three novel conopeptides named as Vi14b, Vi002, Vi003, three conotoxin variants named as Mr3d.1, Mr3e.1, Tx3a.1, and three known conotoxins (Vi15a, Mr3.8 and TCP) from crude venoms of Conus virgo, Conus marmoreus and Conus texile collected in the South China Sea. In this work, we reported their isolation, sequence identification, synthesis, disulfide connectivity recognition and biological evaluation.
2. Results and Discussions
2.1. Peptide Isolation and Sequencing
Crude venom powders weighing 4.1 mg, 18.9 mg and 11.6 mg were prepared from the venom dusts of C. virgo, C. marmoreus and C. texile snail specimens, respectively. Through systemic separation, three novel conopeptides named as Vi14b, Vi002, Vi003, three conotoxin variants named as Mr3d.1, Mr3e.1, Tx3a.1, and three known conotoxins (Vi15a, Mr3.8 and TCP) were isolated and identified from the crude venoms of these three species of cone snails (Table 1).
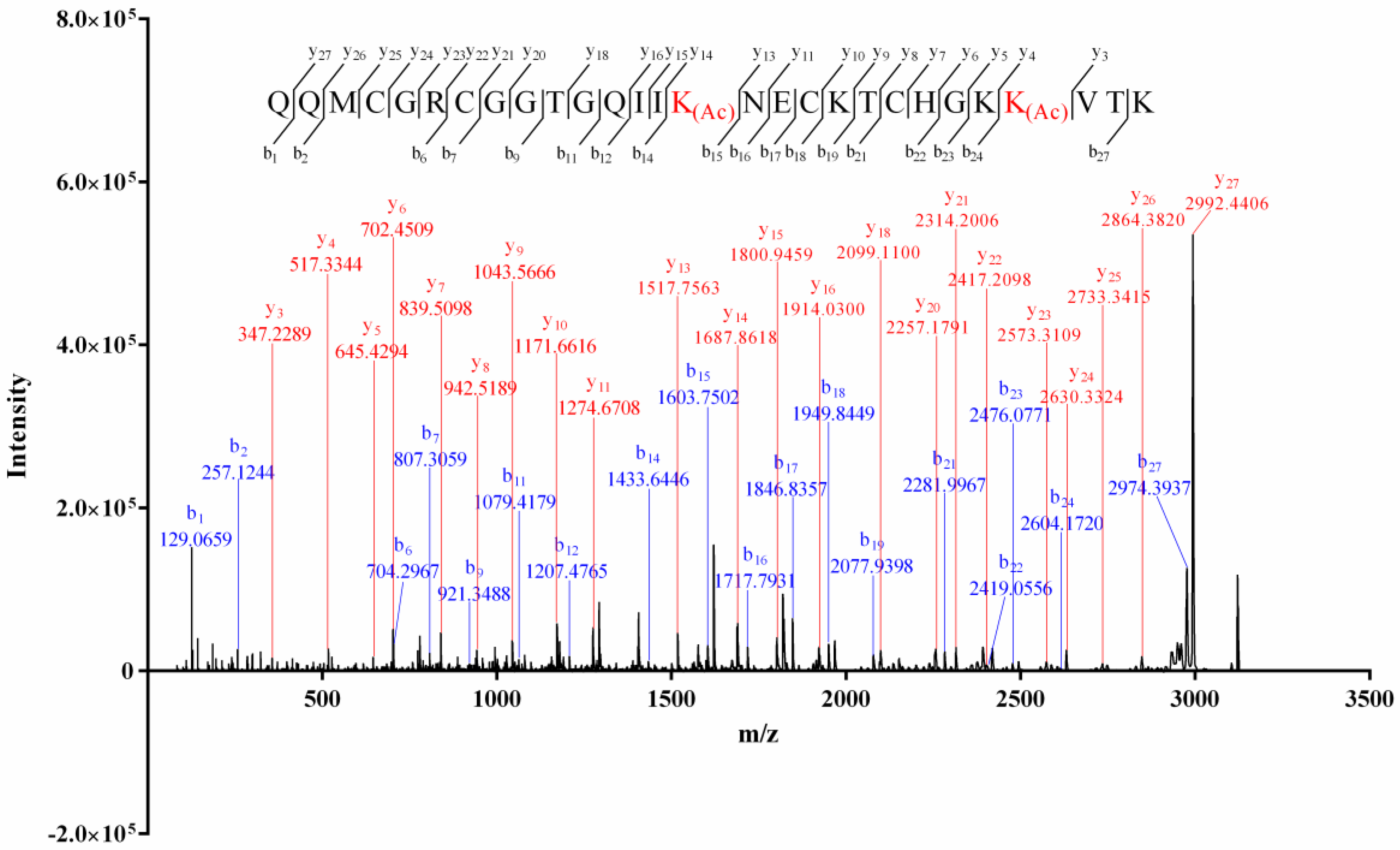
The alkylation result of the isolated conopeptides showed that none of them contained free thiol. The mass difference (4.03 Da, Supplemental Material A, Figure S1) between the intact and the reduced Vi14b indicated that it contained two pairs of disulfide bridges. The Edman degradation result (Supplemental Material B, Figure S2) indicated a sequence of QQM*GR*GGTGQII*NE*KT*HGK*V**. The matrix-assisted laser desorption-ionization–time of flight-tandem mass (MALDI-TOF-MS/MS) spectrum of Vi14b was shown in Figure 1. The serial b1–b2, b6–b7, b9, b11–b12, b14, b15–b19, b21–b24, b27, y3–y11, y13–y16, y18, y20–y27 ions, combined with the key fragment ions (Table 2), indicated that its primary sequence was QQMCGRCGGTGQIIK(Ac)NECKTCHGKK(Ac)VTK, which corresponded with the Edman degradation data. This novel sequence was named Vi14b, since it was the second conotoxin discovered from C. virgo whose cysteine arrangement pattern (C-C-C-C) was framework XIV. Thereinto, two lysines (Lys, K) at the positions 15 and 25 of Vi14b were acylated since the mass addition of Lys was 42.0319, while the Lys at the positions 19, 24 and 28 were not modified. Acylation of lysine is a common PTM (posttranslational modification) among conotoxins.
Conotoxins Vi002 and Vi003 both contained no disulfide bridge since their molecular weights before and after TCEP reduction were the same. In MALDI–TOF–MS/MS spectrum of Vi002 (Figure 2), the consecutive ions b1–b3, a1–a9, y13–y15 and the crucial ions (b10, b14, b17, b21, y1, y3, y5, a17, a20, a22, a24), integrating with the key fragment ions (Table 3), explicitly revealed a novel sequence of LSSGATALSGVPRLTKPAGRLTTTTVAVAF. For sequence identification of Vi003, the serial b1–b7, b9–b11, y3–y4, y7–y9, y11 ions and the key fragment ions (KG m/z 186.1237, KGE m/z 287.1714, KGES m/z 402.1983, KGESL m/z 515.2824, KEGSLL m/z 628.3665, KEGSLLG m/z 685.3879) were observed in the tandem mass spectrum (Supplemental Material A, Figure S2), indicating a novel sequence of NTESTKGESLLGK. It was a pity that we did not obtain enough Vi002 and Vi003 for Edman experiments. The amino acid “L” in the sequences of Vi002 and Vi003 represented “I/L”, since MS/MS sequencing could not distinguish leucine (L) and isoleucine (I). Herein, primary sequences of Vi002 and Vi003 were temporarily determined to be (I/L)SSGATA(I/L)SGVPR(I/L)TKPAGR(I/L)TTTTVAVAF and NTESTKGES(I/L)(I/L)GK, respectively.
Vi15a contained four pairs of disulfide bonds since the mass addition after the TCEP reduction was 8 Da (Figure S1 in Supplementary Materials A). The MS/MS spectrum (Figure S3 in Supplementary Materials A) showed successive ions b1–b8, b13–b16, b18–b22, b24–b26, a22–a26, y15–y26, y1–y3 and the key fragment ions (TCP m/z 302.1169, TCPW m/z 488.1962, TCPWG m/z 545.2177, DNCS m/z 420.1184, DNCSC m/z 523.1275, DNCSCI m/z 636.2116), which precisely determined the sequence to be DCTTCAGEECCGRCTCPWGDNCSCTEW-(nh2), a previously found disulfide-rich conotoxin Vi15a from C. virgo [24].
The mass differences between the reduced and unreduced conopeptides Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 and TCP were 6 Da (Figure S1 in Supplementary Materials A), respectively, indicating that these five conotoxins all contained three pairs of disulfide bridges. The secondary MS spectra of the five conopeptides were shown in Figures S4–S8 of Supplemental Material A, while their Edman degradation results were exhibited in Figures S3–S6 of Supplemental Material B. The sequential ions b2–b13 observed in the MS/MS spectrum (Figure S4 in Supplementary Materials A), combined with the deduced sequence of **RLS*GL**HP** from the Edman degradation results (Figure S3 in Supplementary Materials B), ascertained a novel sequence of CCRLSCGLGCHPCC named as Mr3d.1. The Proline at the position 12 of Mr3d.1 was unmodified, while the 12-Proline of a known conotoxin Mr3d (CCRLSCGLGCHOCC-nh2, O presents “4-hydroxyproline”) [26,28] was hydroxylated and its C-terminal was amidated. Mr3e.1 was identified to be VCCPFGGCHELCYCCD since the consecutive ions b2–b5, b7–b15 were detected in MS/MS spectrum (Figure S5 in Supplementary Materials A), and the Edman degradation result (Figure S4 in Supplementary Materials B) showed a sequence of V**PFGG*HEL*Y***. Herein, Mr3e.1 was a C-terminal deamidated variant of conotoxin Mr3e (VCCPFGGCHELCYCCD-nh2, [25,26,29]). Mr3.8 (CCHWNWCDHLCSCCGS) was a known conotoxin elucidated by serial ions b3–b15, y8–y14 in MS/MS data (Figure S6 in Supplementary Materials A) and the Edman degradation experiment (**HWNW*DHL*S****, Figure S5 in Supplementary Materials B). In Figure S7 in Supplementary Materials A, the serial y7–y9, y11–y15, b5–b6 and b8–b15 ions clearly confirmed the sequence to be CCSWDVCDHPSCTCCG, which was a C-terminal carboxylated variant of a known conotoxin Tx3a (CCSWDVCDHPSCTCCG-nh2) from C. texile [27,28]. Thus, it was named Tx3a.1. The sequence for Figure S8 in Supplementary Materials A was assigned NCPYCVVYCCPPAYCEASGCRPP (a previously found conotoxin named TCP [27]) by successive b/y ions (y1–y22, b1–b3, b5–b9, b11, b21–b22). The Edman degradation result (N*PY*VVY***PAY*E*SG****, Figure S6 in Supplementary Materials B) also supported the above sequence deduced from MS/MS sequencing.
The existence of disulfide bonds is of great significance for stabilizing conotoxins’ spatial structures, enhancing activity and improving selectivity to target receptors [30]. Different disulfide linkages (quantity or connectivity pattern) produce different three-dimensional structures, which lead to differences in activity and selectivity afterwards [30,31,32]. In this work, we isolated a novel two-disulfide-bonded conotoxin Vi14b, two novel disulfide-poor conotoxins Vi002 and Vi003, and five three-disulfide-bridged conotoxins Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 and TCP. Conotoxins Mr3.8 and TCP were previously discovered from Conus venom isolation or transcriptomic/proteomic analyses [25,26,27,28,33]. C.D. Poulter et al. synthesized Tx3a and characterized its disulfide connectivity as I-V, II-IV, III-VI. They found that Tx3a can lead to an excitation response by injecting it into mice [34]. TCP (textile convulsant peptide) was an O-superfamily convulsant peptide with a VI/VII Cys framework purified from C. texitle venom components that had induced dramatic and diverse symptoms in mice [28,35,36]. To date, no other record of biological activity and target for Mr3.8, Tx3a.1 and TCP has been reported.
Since the amount of the isolated peptides in this work was quite limited, in order to investigate the structure and activity of these disulfide-rich conotoxins, we synthesized conotoxins Vi14b, Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 and TCP using a one-step oxidation strategy. For every conotoxin, we individually performed the HPLC co-elution of the isolated peptide and its oxidative products in order to track the desired isomer that was consistent with the native peptide. Subsequently, the correct isomer was purified by HPLC system. The homogeneities between the synthetic peptide and the corresponding isolated one were verified by the HPLC co-elution experiment.
2.2. Disulfide Connectivity
Conotoxins Vi14b, Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 and TCP were synthesized using a one-step oxidation strategy. The disulfide connectivities of these folded peptides were unknown. Herein, their disulfide linkages were determined by MALDI–TOF–MS/MS analysis of their two-cysteine-alkylated products (product I) and the four-cysteine-alkylated products (product II) generated from the stepwise reduction and subsequent alkylation of these folded peptides (Figure 3 and Figure S9 in Supplementary Materials A).
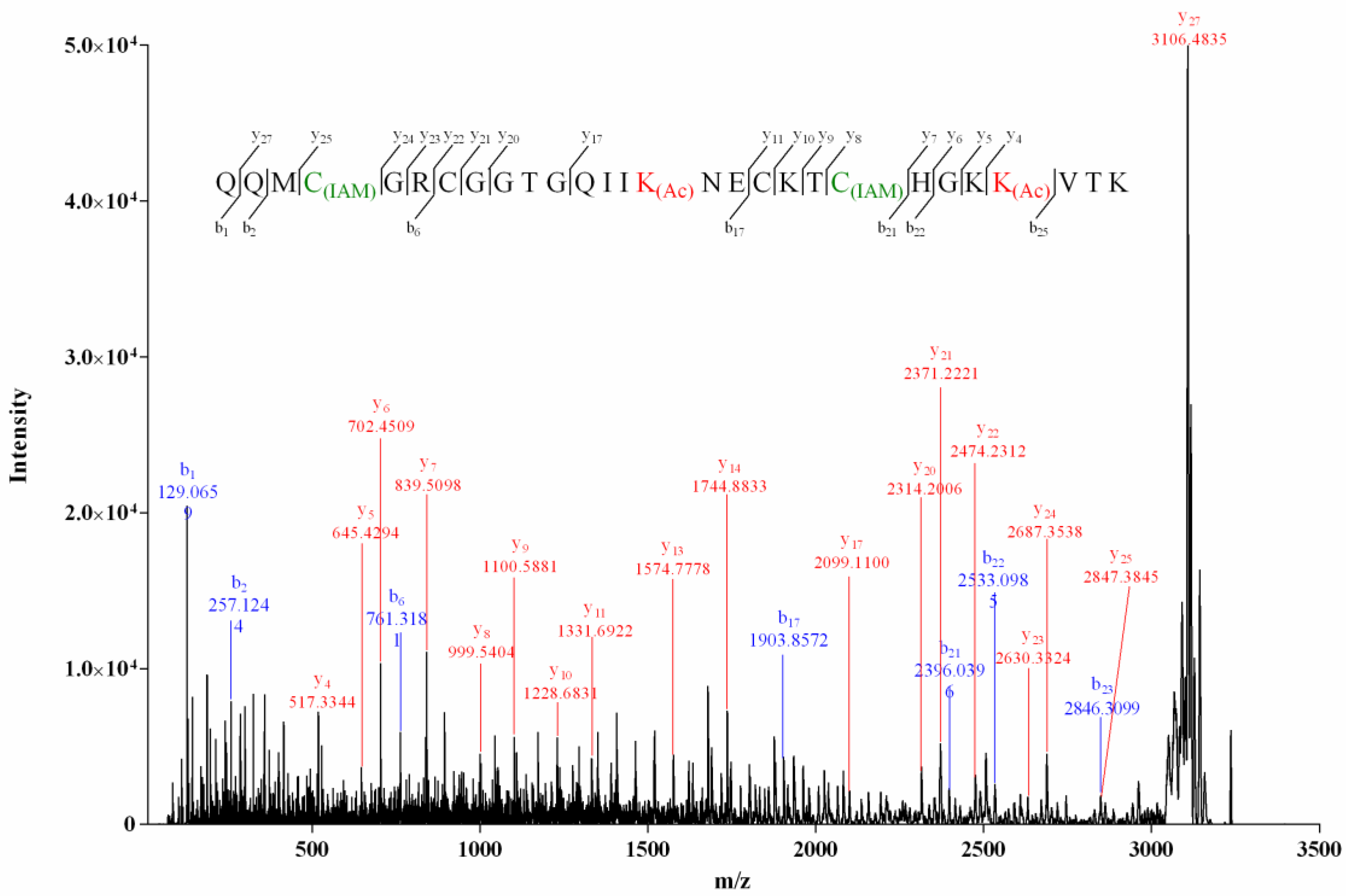
The MS/MS spectrum of the reduced two-cysteine-alkylated product (product I, Figure 3) of Vi14b was shown in Figure 4. We used “C(IAM)” to represent the cysteine that was alkylated by IAM (Iodoacetamide) in Figure 4, Figure 5 and Figure 6 and Figures S10–S16 in Supplementary Materials A. The mass differences of y25/y24 and y8/y7 were 160.0307 and 160.0306, respectively, while mass differences of y22/y21 and y11/y10 were both 103.0091, indicating that Cys-I and Cys-IV were alkylated while Cys-II and Cys-III were in their reduced form. It meant that a disulfide linkage existed between Cys-I and Cys-IV. Herein, the disulfide connectivity of Vi14b was characterized as I-IV, II-III.
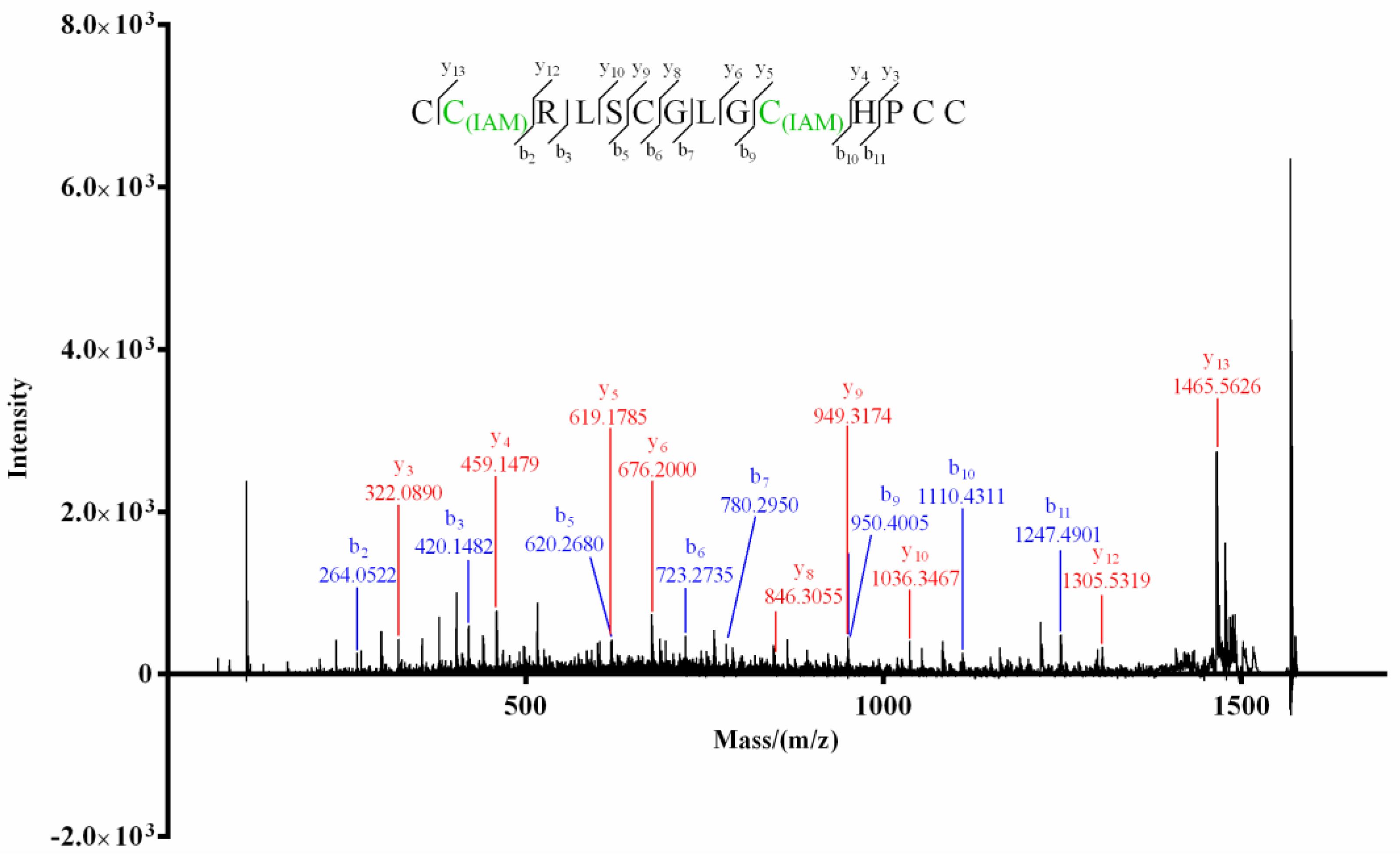
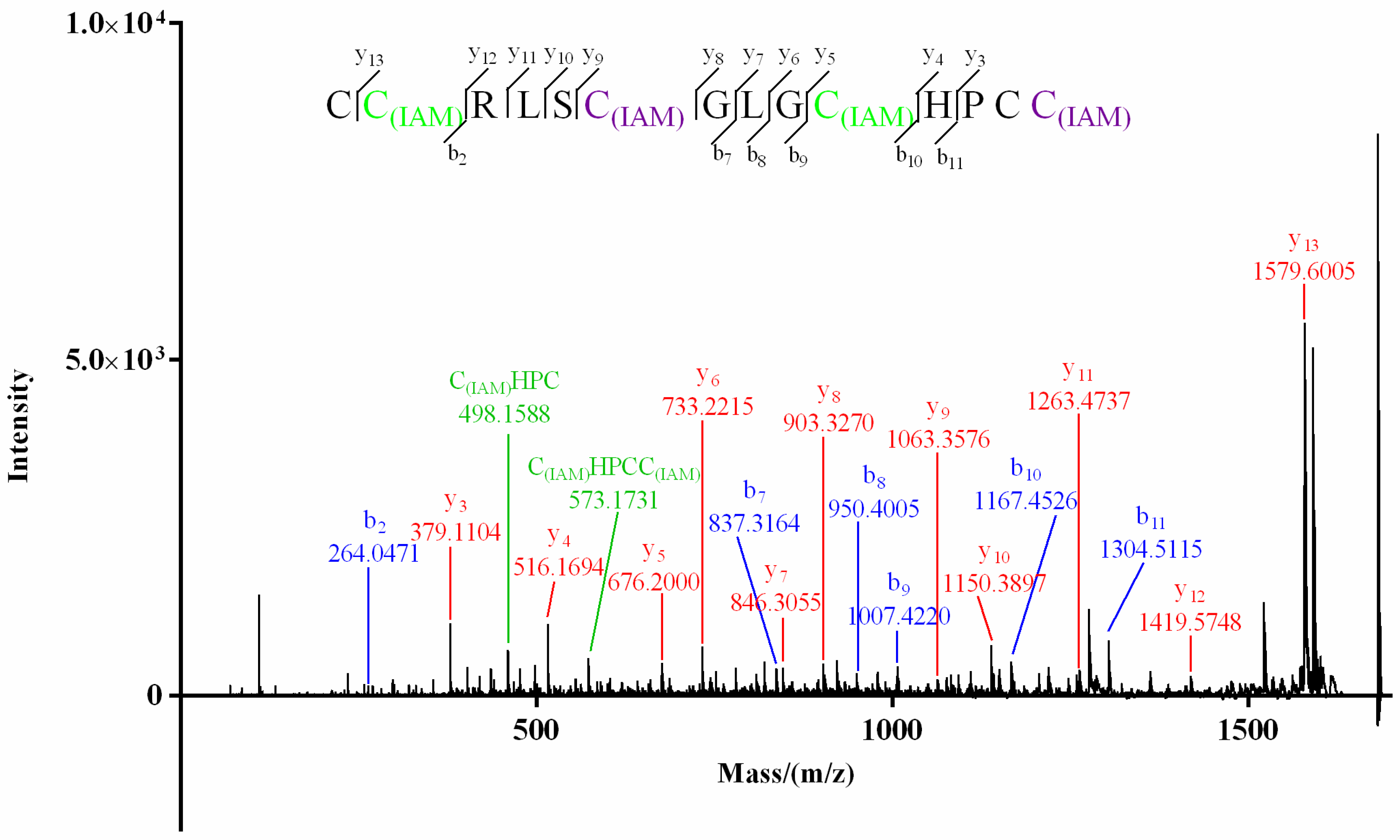
For product I (Figure 3) of Mr3d.1, Cys at positions 2 and 10 (2-Cys and 10-Cys) were speculated to be IAM-labeled since the mass difference (160.0307) of y13/y12 and b10/b9 (or y5/y4) was deduced from its MS/MS spectrum (Figure 5). In the MS/MS spectrum of product II of Mr3d.1 (Figure 6), the mass difference (160.0307) of y13/y12, y9/y8, y5/y4 (or b10/b9) meant the Cys at positions 2, 7 and 10 were alkylated. The observation of ions y14/y13 and fragment ions C(IAM)HPC (m/z 498.1588) and C(IAM)HPCC(IAM) (m/z 573.1731) suggested that thiols in 1-Cys and 13-Cys were in reducing state and 14-Cys were alkylated. Herein, the disulfide linkage of Mr3d.1 was confirmed to be I-V, II-IV, III-VI.
As for product I of Mr3e.1 (Figure S10 in Supplementary Materials A), 3-Cys and 14-Cys were ethyl-labeled because of the mass difference (160.0307) of b3/b2 (or y14/y13) and b14/b13, while the other four cysteines were dissociative. In MS/MS spectrum of product II of Mr3e.1 (Figure S11 in Supplementary Materials A), the ions of y15/y15, b3/b2, b8/b7, b14/b13 revealed the alkylation of Cys at positions 2, 3, 8 and 14. Thus, Mr3e.1 was considered to have I-III, II-V, IV-VI disulfide connectivity. Similarly, for Mr3.8, Cys-II and Cys-VI of its product I (Figure S12 in Supplementary Materials A) were alkylated by the observation of y15/y14 and b14/b13. In MS/MS spectrum of product II of Mr3.8 (Figure S13 in Supplementary Materials A), the detection of ions b14 (m/z 458.1275) and y3 (m/z 323.1020) indicated that the cysteins at positions 1, 2 and 14 were IAM-labeled, while 2-Cys and 10-Cys were in the reducing state due to the mass difference (103.009) of b7/b6 and b11/b10. The 13-Cys were alkylated since ya ion (m/z 573.2272) of the fragment HLCSC(IAM) was detected. Thus, disulfide linkage of Mr3.8 was identified as I-V, II-VI, III-IV.
With regard to Tx3a.1, Cys-I and Cys-III were ascertained to be alkyl-marked in its product I due to the mass difference (160.03) of ions of y10/y9 and y15/parent ion (m/z 1832.5632) (Figure S14 in Supplementary Materials A), while ions of y15/y14, b12/b11, b14/b13, b15/b14 revealed Cys at positions 2, 12, 14 and 15 were in their reduced form. For product II of Tx3a.1 (Figure S15 in Supplementary Materials A), the ions b2 (m/z 321.0608), y2 (m/z 236.0700) and y10/y9 (or b7/b6) ascertained 1-, 2-, 7-, 15-Cys were alkyl-labeled. Thus, the disulfide connectivity of Tx3a.1 was assigned as I-III, II-VI, IV-V. Similarly, disulfide linkages of TCP were notarized as I-III, II-V, IV-VI because of ions of b12/b11 and y15/y14 were observed in the MS/MS spectrum of its product I (Figure S16 in Supplementary Materials A) and ions of b2 (m/z 275.0809), y19/y18, y15/y14, y9/y8 were detected in MS/MS spectrum of its product II (Figure S17 in Supplementary Materials A).
Thus, the disulfide connectivities of conotoxins Vi14b, Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 and TCP were assigned as shown in Figure 7. Generally, linkage patterns of I-V, II-IV, III-VI and I-III, II-V, IV-VI are common among the conotoxins with three pairs of disulfide bonds, while the patterns of I-V, II-VI, III-IV (such as Mr3.8) and I-III, II-VI, IV-V (such as Tx3a.1) were rarely reported and different from that previously established for the µ- and ψ-conotoxins [37]. The novel connectivity patterns could possibly cause these kinds of conotoxins, such as Tx3a.1 and Mr3.8, to present specific biological activity.
2.3. Blockage on Voltage-Gated Sodium Channel
The Cys arrangement pattern of Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 was framework III, while that of TCP was framework VI/VII. Frameworks III and VI/VII are common in μ-conotoxins, which generally target voltage-gated sodium channels (VGSC). Sodium channel subtypes can be classified as TTX-sensitive (Nav1.1, 1.2, 1.3, 1.4, 1.6 and 1.7), or TTX-resistant (Nav1.5, 1.8, 1.9) channels upon their sensitivity to tetrodotoxin (TTX) [13]. Nav1.4 is highly expressed in adult skeletal muscle and is associated with disorders such as myotonia, myasthenia and periodic paralysis [23]. A potent and selective Nav1.4 blocker μ-CnIIIC (IC50 1.3 nM, framework III) has been commercialized as an instant line relaxer called XEPTM-018 and been used in a facial cream (LirikosTM, Amorepacific Corporation) for wrinkle smoothing [38]. Nav1.8 is generally distributed in DRG (dorsal root ganglion) neurons and is considered to be an important target for chronic pain therapy [39]. μO-MrVIB (IC50 326 nM for Nav1.8, framework VI/VII) is a potential drug for neuropathic pain treatment, and its investigation has entered clinical phase II [40]. Here, we evaluated the effect of conotoxins Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1, TCP on Nav1.4 and Nav1.8 channels. The result (Table 4 and Figure 8) indicated that conotoxins Mr3e.1, Mr3.8, Tx3a.1, and TCP blocked Nav1.4 or Nav1.8 by slight inhibition on the current at the complete resting state (TP1), while they inhibited Nav1.4 or Nav1.8 by 21.51~24.32% of currents in a semi-activated state (TP2) at 10 μmol/L. Other tested conotoxins showed no current inhibition on Nav1.4 or Nav1.8. Activities on other subtypes of VGSC or other types of ion channels remain to be determined.
2.4. Tx3a.1, Mr3.8 and Vi14b Inhibited the Proliferation of ID8 Cells
To study the anticancer potential of conotoxins Vi14b, Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1, TCP, the cytotoxicity of these peptides against ID8 ovarian cancer cells was detected by MTT assay. We treated the cells with different concentrations (0, 5, 10, 20, 40, 80, 160 μM) of peptides for 72 h. These peptides showed significant inhibitory effect on ID8 cells at a low concentration of 5 μM compared with the control group, especially Tx3a.1. The cell inhibitory rates were 45.08%, 23.79% and 19.39% for Tx3a.1, Mr3.8 and Vi14b, respectively, at a concentration of 10 μM. A stronger effect was observed after exposure to higher concentrations of Tx3a.1, with a 50% reduction in cell viability at a concentration of 24.29 μM. Mr3.8 killed half the cells at a concentration of 54.97 μM. The IC50 of Vi14b was 111.6 μM. The data (Figure 9) were presented as mean ± SD of three independent experiments. These results indicated that the Tx3a.1, Mr3.8 and Vi14b exhibited strong anticancer effects on ID8 cells in vitro. It was the first report of conotoxins showing anti-ovarian cancer activity. The mechanism of their anti-ovarian cancer activity remains to be explored later. In this work, we identified and structurally and biologically characterized the anti-ovarian cancer conotoxins from Conus venom. The result delivered a message that conotoxins, a class of active polypeptides with diverse structures, could be a rich source of anti-ovarian cancer drug candidates. Herein, this study highlights the role of conotoxin library in subsequent drug discovery for ovarian cancer treatment.
3. Materials and Methods
3.1. Venom Preparation and Peptide Isolation
Cone snail specimens were collected from coastal waters near Sansha city in the South China Sea and were frozen at −80 °C for hours. Dr. Ying Fu identified the specimens. The venom duct of the snails was dissected, and the venom duct cut into small sections (about 1cm long). They were extracted with 60% acetonitrile aqueous solution at 4 °C for 24 h, then centrifuged at 8000 r/min for 10 min. The supernatant was pipetted and filtered with 0.22 µm organic phase pinhole filter, then 60% acetonitrile aqueous solution was added to the precipitate for secondary extraction. This was repeated twice and the filtrate combined to obtain the crude venom solution, which was subjected to lyophilization to yield crude venom powder.
The venom powder was dissolved in 3% acetonitrile aqueous solution and filtrated. The venom solution was fractionated using a preparative high-performance liquid chromatography (Pre-HPLC) instrument (Waters, Milford, MA, USA) with a C18 column (Vydac Grace, 10 µm, 22 mm × 250 mm). The preparative system was conducted by linear gradient elution with 5 to 90% solution B (90% acetonitrile aqueous solution with 0.1% TFA) for 60 min at an 8 mL/min flow rate, while solution A was double-distilled water (ddH2O) with 0.1% TFA. The fractionated components were lyophilized and redissolved in 10% acetonitrile aqueous solution, then were subjected to a HPLC system equipped with a C18 column (Vydac Grace, 5 µm, 4.6 mm × 250 mm). The separation process was performed with mixed eluant (solution A and B) at a flow rate of 0.8 mL/min. Conopeptides were yielded from the venom fractions by a 20-min linear gradient program of 30% to 50% solution B (namely 70% to 50% solution A) detecting by ultraviolet (UV) absorption at 214 nm.
The molecular weights of the purified conopeptides were determined using an ultra-performance liquid chromatography–triple quadrupole-mass spectrometry (UPLC–TQD–MS, Waters, USA) spectrometer with a BEH C18 column (130 A, 1.7 µm, 2.1 mm × 100 mm). Conditions were set as follows: detection range m/z 400–1500, cone voltage 30 V, capillary voltage 3.5 kV, dissolution flow rate 1000 L/h, dissolution temperature 500 °C, ion source temperature 150 °C. The UPLC program was performed by gradient elution from 5 to 60% solution B for 6 min at a flow rate of 0.5 mL/min.
3.2. Peptide Sequencing
The isolated peptides were individually alkylated by iodoacetamide to confirm whether they contained any free thiol or not. In order to ascertain the number of disulfide bonds in these conopeptides, we totally reduced the peptides by TCEP and detected their mass differences between intact peptides and the reduced forms (Figure 3), respectively.
Then the purified conotoxins were totally reduced to obtain linear peptides before sequencing. Each peptide was dissolved in 20 µL 50% acetonitrile aqueous solution. Then, 10 μL 50 mmol/L TCEP and 10 μL NH4HCO3 were added to the peptide solution, which was incubated at 25 °C for 30 min. The reaction was monitored by UPLC analysis at 214 nm. The molecular weight of each reduced peptide was determined by UPLC–TQD–MS over m/z 100–2000.
Sequence identification was achieved by MALDI–TOF (matrix-assisted laser desorption—ionization—time of flight) tandem mass spectrometer (Bruker, Ultraflextreme, Germany). The reduced linear peptides were dissolved in 50% acetonitrile aqueous solution. The matrix solution was prepared by dissolving 1 mg cyano-4-hydroxycinnamic acid (HCCA) in 100 µL standard solution (47.5 µL ddH2O, 50 µL acetonitrile, 2.5 µL trifluoroacetic acid). For sample loading, 1 µL of each reduced peptide solution was individually dropped onto different circles on the plate and then dried off. One µL of matrix solution was successively dropped onto the same position on the plate. The sample plate was then loaded into the spectrometer when the solution was completely dried off. FlexControl was used to acquire MS data, while FlexAnalysis was applied to mark the masses. Conditions were set as follows: laser frequency 1000 Hz, laser energy range 20–40%, ion source voltage 7.5 kV. The voltages for secondary mass detection were 19.00 kV for Lift 1 and 2.80 kV for Lift 2. For the sequence identification, ion masses and related fragmentation masses in the secondary mass spectrum of each conopeptide were matched by Mascot search (http://www.matrixscience.com/cgi/nph-mascot.exe?1, accessed from 1 December 2020 to 23 February 2022) against the NCBI and Swiss-prot protein database. Modifications involved in the search were as follows: amidation, deamidation, hydroxylation of proline and valine, oxidation of methionine, carboxylation of glutamic acid, cyclization of N-terminal glutamine (pyroglutamate), bromination of tryptophan, and sulfation of tyrosine.
An Edman degradation experiment was performed for sequence elucidation as well. The solution was vortexed until the reduced peptide powder was dissolved in 50 μL ddH2O. Then, 10 μL of the peptide solution was dropped onto the polyvinylidene fluoride (PVDF) membrane and dried off. The PVDF membrane was transferred onto a PPSQ-53A protein sequencer (Shimadzu, Japan). Each phenylthiohydantoin (PTH)-amino acid generated from every cycle of N-terminal degradation of the peptide was analyzed by HPLC system equipped with a Wakosil PTH-II column (Wako, 4.6 mm × 250 mm). For the mobile phase, 40% acetonitrile aqueous solution with less than 2.5% acetic acid was used. The column temperature was set to 40 °C, and the flow rate was 1.0 mL/min. The retention time of each (PTH)-amino acid was compared with that of the corresponding standard amino acid to identify the primary amino acid sequence of the isolated conopeptides.
3.3. Peptide Synthesis
The linear conopeptides were separately synthesized using Fmoc-SPPS (9-Fluorenylmethoxycarbonyl-peptide solid phase synthesis) chemistry [41]. The Fmoc-amino acids were purchased from Chengdu Chron Chemical Co., LTD. Amino acid ligation was operated on the Wang-resin with HBTU/DIEA used as the coupling reagent. Kaiser test regent was applied to detect the state of every coupling cycle. Piperidine/DMF (v/v 20:80) was used to remove the Fmoc groups. The cysteine residues were protected by triphenylmethyl (trt) groups, which were then removed along with the peptide release process. Cleavage cocktail (TFA/thioanisole/phenol/EDT/H2O 87.5:2.5:2.5:2.5) was used to release the linear peptide from the resin at room temperature for 2 h, filtrate to remove the resin, and the filtrate was treated with cold ether (0 °C) overnight to precipitate the crude peptide. The precipitation was washed with cold ether twice and lyophilized to obtain crude peptide power, which was subjected to Pre-HPLC separation to yield pure linear peptide.
The obtained linear conopeptide was subjected to one-step air oxidation in a 0.1 M Tris-HCl/1 mM EDTA buffer with GSH/GSSG (1 mmol/L/0.5 mmol/L) at 40 °C for 60 min. The linear peptide concentration ranged from 50 μM to 100 μM. The oxidative products were detected by LC–MS and subsequently purified by HPLC system with a 20-min linear gradient elution of 30% to 50% solution B at a 0.8 mL/min flow rate.
3.4. Disulfide Connectivity Recognition
The stepwise reduction and alkylation protocol were performed as reported [42]. The folded peptide (20–30 μg in 100 μL ddH2O containing 0.1% TFA) was partially reduced by mixing with an appropriate volume of 100 mM TCEP dissolved in citrate buffer (0.1 M, pH 3.0) to a final concentration of 6 mM TCEP. The mixture was incubated at 40 °C ranging from 5 min to 25 min for folded peptide to generate the corresponding one-disulfide-reduced product and two-disulfide-reduced product. The reaction state was monitored by LC–MS to confirm that the exact partially reduced product was generated. Then the reaction mixture was blended with alkylation solution (9.24 mg iodoacetamide dissolved in 25 μL acetonitrile and 75 μL 1 mM pH 8.0 Tris-HCl buffer) to alkylate the free thiol groups of the partial-reduced products. The alkylation reaction was conducted at 40 °C for 15–45 min and monitored by LC-MS. Then the mixture was subjected to going through HPLC column (Vydac Grace, 5 μm, 4.6 mm × 250 mm) to terminate the reaction and purify the alkyl-labeled products. The obtained two-cysteine-alkylated and four-cysteine-alkylated products were subsequently reduced by TCEP to break the remaining disulfide bonds to gain linear peptides, which were named as products I and II, respectively. Then the products I and II of the synthetic conopeptide were subjected to MALDI–TOF–MS/MS analysis to confirm which thiol groups were alkylated.
3.5. Patch Clamp Recording for Voltage-Gated Sodium Channel
Blocking activity of voltage-gated sodium channels was conducted using the whole-cell patch clamp technique [43]. CHO cells were cultured in HAM’S/F-12 medium containing 10% fetal bovine serum. 6.5 × 103 cells were plated on coverslips, cultured in 24-well plates (final volume 500 µL). Patch clamp assays were performed after 18 h. Solution preparation involved intracellular fluid Nav-001-2 (50 mM CsCl, 10 mM NaCl, 10 mM HEPES, 60 mM CsF, 20 mM EGTA, adjusted to pH 7.2 with CsOH), extracellular fluid K-007-1 (140 mM NaCl, 3.5 mM KCl, 1 mM MgCl2•6H2O, 2 mM CaCl2•2H2O, 10 mM D-Glucose, 10 mM HEPES, 1.25 mM NaH2PO4•2H2O, adjusted to pH 7.4 with NaOH). The testing peptide was dissolved with DMSO to a concentration of 10 mM solution. The peptide solution was diluted to 10 µM using extracellular fluid. Tetrodotoxin (TTX, 1 µM) dissolved in ddH2O containing 0.1% acetic acid was used as a Nav1.4 positive control, while Nav1.8 positive control was prepared by dissolving 10 mg A-803467 [5-(4-chlorophenyl-N-(3, 5-dimethoxyphenyl) furan-2-carboxamide] with DMSO and diluting it to 10 µM by K-007-1). The microelectrode manipulator was operated under an inverted microscope (MP285, Sutter Instrument, Novato, CA, USA). The recording electrode was placed on the cell, and negative pressure was applied to form a GΩ seal. Then, fast capacitance compensation was performed and negative pressure applied until the cell membrane was sucked to form a whole-cell recording mode. The membrane voltage was clamped at −120 mV after whole-cell sealing. Cell-plated coverslips were placed in the recording bath in an inverted microscope. The testing peptide solution and the peptide-free extracellular fluid flowed through the recording bath from low to high concentration by gravity perfusion. Each peptide was reacted for 5 min before the next peptide was detected. The data were collected by EPC-10 amplifier (HEKA, Reutlingen, Baden Wurdenburg, Germany) and stored by PatchMaster software (HEKA, Reutlingen, Baden Wurdenburg, Germany).
The reaction rate on voltage-gated sodium channel were defined as , and the percentage of inhibition was determined as 1 − × 100%. Meanwhile, the mean and standard deviation were calculated through three parallel experiments. The voltage-gated sodium channel activity data was analyzed by PatchMaster (HEKA, Reutlingen, Germany) & IGOR Pro (Wave Metrics, Portland, OR, USA). In addition, data processing and graphing were performed by the software GraphPad Prism 6.0 (GraphPad Software, San Diego, CA, USA).
3.6. Ovarian Cancer Cell Viability Assay
The MTT method was used to determine the cytotoxicity of the synthetic conotoxins [44]. Briefly, ID8 cells (4 × 103 cells/well) were seeded in 96-well plates. After 12 h incubation, the culture medium was removed and the cells were treated with different concentrations of peptides for 72 h. After adding 1 mg/mL MTT solution, the cells were cultured for another 4 h. Subsequently, 100 µL DMSO was added to dissolve the formazan. The absorbance was measured at 570 nm using a SpectraMax® M2 microplate reader (Molecular Devices, Sunnyvale, CA, USA). The IC50 values were expressed as mean ± SD of individual samples and were representative of three independent experiments. The data were analyzed by unpaired two-tail t-test (Student’s t-test). Values of P less than 0.05 were regarded as statistically significant. Calculation of IC50 values and statistical analysis were performed using GraphPad Prism 6.0 (GraphPad Software, San Diego, CA, USA).
4. Conclusions
We isolated and identified three novel conopeptides named as Vi14b, Vi002, Vi003, three conotoxin variants named as Mr3d.1, Mr3e.1, Tx3a.1, and three known conotoxins (Vi15a, Mr3.8 and TCP) from Conus venom. Mr3.8 and Tx3a.1 both showed a novel pattern of disulfide connectivity, different from that previously established in the µ- and ψ-conotoxins. Tx3a.1, Mr3e.1 and Vi14b exhibited a certain anti-ovarian cancer effect on ID-8 cells in vitro. It was the first report of conotoxins showing anti-ovarian cancer activity. This work highlights the role of conotoxin library in subsequent drug discovery for ovarian cancer treatment.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules27196609/s1. HPLC chromatograms, primary and secondary mass spectra (Figures S1–S17 (Supplementary Materials A)) that were mentioned in this article are shown in Supplementary Material A. Edman degradation data (Figures S1–S6 and Table S1 (Supplementary Materials B)) were exhibited in Supplementary Material B.
Author Contributions
Conceptualization, S.L. (Sulan Luo) and Y.F.; Data curation, S.J., Y.Z., X.G. and B.M.; Formal analysis, S.J., Y.Z., X.G. and B.M.; Funding acquisition, J.B., S.L. (Sulan Luo) and Y.F.; Investigation, S.J., Y.Z. and X.G.; Methodology, Y.Z., Q.Y. and S.L. (Siyi Liu); Project administration, S.L. (Sulan Luo) and Y.F.; Resources, J.B., S.L. (Sulan Luo) and Y.F.; Supervision, S.L. (Sulan Luo) and Y.F.; Validation, Q.Y., S.L. (Siyi Liu) and M.Z.; Writing—original draft, S.J., X.G. and Y.F.; Writing—review & editing, J.B. and Y.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hainan Provincial Natural Science Foundation of China (No. 2019RC053) and National Natural Science Foundation of China (No. 81903492 and No. 82104057).
Institutional Review Board Statement
The animal study protocol for X. Laevis was approved by the Ethics Committee of Hainan University (protocol code IACUC-HaiDaBan-2020-17 and 13 April 2020 of approval).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
DMF: dimethyl formamide; DMSO: dimethyl sulfoxide; EDTA: ethylenediaminetetraacetic acid; ESI–TQD: electrospray ionization-triple quadrupole; GSH: glutathione; GSSH: glutathione (oxidized); HPLC: high performance liquid chromatography; IAM: Iodoacetamide; MALDI–-TOF–MS/MS: matrix-assisted laser desorption ionization-time of flight-tandem mass; PARP: poly ADP-ribose polymerase; PTM: posttranslational modification; TCEP: tris-(2-carboxyethyl)phosphine hydrochloride; TFA: trifluoroacetic acid; Tris-HCl: trihydroxymethylaminomethane hydrochloride; VGSC: voltage-gated sodium channels; UPLC: Ultra performance liquid chromatography.
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.; Ralyea, C.; Lockwood, S. Ovarian Cancer: An Integrated Review. Semin. Oncol. Nurs. 2019, 35, 151–156. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Surveillance, Epidemiology, and End Results Program. Cancer Stat Facts: Ovarian Cancer. Available online: https://seer.cancer.gov/statfacts/html/ovary.html (accessed on 24 August 2022).
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; Ledermann, J.A. Ovarian cancer. Lancet 2014, 384, 1376–1388. [Google Scholar] [CrossRef]
- Vargas, A.N. Natural history of ovarian cancer. Ecancermedicalscience 2014, 25, 465–476. [Google Scholar]
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [Green Version]
- Cortez, A.J.; Tudrej, P.; Kujawa, K.A.; Lisowska, K.M. Advances in ovarian cancer therapy. Cancer Chemother. Pharmacol. 2018, 81, 17–38. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, D.M. New Therapies for Ovarian Cancer. J. Natl. Compr. Canc. Netw. 2019, 17, 619–621. [Google Scholar]
- Boyd, L.R.; Muggia, F.M. Carboplatin/Paclitaxel Induction in Ovarian Cancer: The Finer Points. Oncology 2018, 32, 418–420, 422–424. [Google Scholar]
- Buczek, O.; Bulaj, G.; Olivera, B.M. Conotoxins and the posttranslational modification of secreted gene products. Cell Mol. Life Sci. 2005, 62, 3067–3079. [Google Scholar] [CrossRef]
- Gao, B.; Peng, C.; Yang, J.; Yi, Y.H.; Zhang, J.Q.; Shi, Q. Cone snails: A big store of conotoxins for novel drug discovery. Toxins 2017, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Antunes, A. Biomedical Potential of the Neglected Molluscivorous and Vermivorous Conus Species. Mar. Drugs. 2022, 20, 105. [Google Scholar] [CrossRef]
- Robinson, S.D.; Norton, R.S. Conotoxin gene superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [Green Version]
- Han, T.S.; Teichert, R.W.; Olivera, B.M.; Bulaj, B. Conus venoms—A rich source of peptide-based therapeutics. Curr. Pharm. Des. 2008, 14, 2462–2479. [Google Scholar] [CrossRef]
- Vetter, I.J.; Lewis, R. Therapeutic potential of cone snail venom peptides (conopeptides). Curr. Top Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef]
- Quik, M.; Wonnacott, S. α6β2* and α4β2* nicotinic acetylcholine receptors as drug targets for Parkinson’s disease. Pharmacol. Rev. 2011, 63, 938–966. [Google Scholar] [CrossRef] [Green Version]
- Becchetti, A.; Aracri, P.; Meneghini, S.; Brusco, S.; Amadeo, A. The role of nicotinic acetylcholine receptors in autosomal dominant nocturnal frontal lobe epilepsy. Front. Physiol. 2015, 6, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef]
- Gallo, A.; Boni, R.; Tosti, E. Neurobiological activity of conotoxins via sodium channel modulation. Toxicon 2020, 187, 47–56. [Google Scholar] [CrossRef]
- Dhiman, V.; Pant, D. Human health and snails. J. Immunoass. Immunochem. 2021, 42, 211–235. [Google Scholar] [CrossRef]
- Fu, Y.; Li, C.; Dong, S.; Wu, Y.; Zhangsun, D.; Luo, S. Discovery Methodology of Novel Conotoxins from Conus Species. Mar. Drugs 2018, 16, 417. [Google Scholar] [CrossRef] [Green Version]
- Pope, J.E.; Deer, T.R. Ziconotide: A clinical update and pharmacologic review. Expert Opin. Pharmacother. 2013, 14, 957–966. [Google Scholar] [CrossRef]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and Biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef]
- Peng, C.; Liu, L.; Shao, X.; Chi, C.; Wang, C. Identification of a novel class of conotoxins defined as V-conotoxins with a unique cysteine pattern and signal peptide sequence. Peptides 2008, 29, 985–991. [Google Scholar] [CrossRef]
- Wang, Q.; Jiang, H.; Han, Y.H.; Yuan, D.D.; Chi, C.W. Two different groups of signal sequence in M-superfamily conotoxins. Toxicon 2008, 51, 813–822. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.H.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [Green Version]
- Tayo, L.L.; Lu, B.; Cruz, L.J.; Yates, J.R., 3rd. Proteomic analysis provides insights on venom processing in Conus textile. J. Proteome Res. 2010, 9, 2292–2301. [Google Scholar] [CrossRef] [Green Version]
- Corpuz, G.P.; Jacobsen, R.B.; Jimenez, E.C.; Watkins, M.; Walker, C.; Colledge, C.; Garrett, J.E.; McDougal, O.; Li, W.; Gray, W.R.; et al. Definition of the M-conotoxin superfamily: Characterization of novel peptides from molluscivorous Conus venoms. Biochemistry 2005, 44, 8176–8186. [Google Scholar] [CrossRef]
- Han, Y.H.; Wang, Q.; Jiang, H.; Liu, L.; Xiao, C.; Yuan, D.D.; Shao, X.X.; Dai, Q.Y.; Cheng, J.S.; Chi, C.W. Characterization of novel M-superfamily conotoxins with new disulfide linkage. FEBS J. 2006, 273, 4972–4982. [Google Scholar] [CrossRef]
- Gngora-Bentez, M.; Tulla-Puche, J.; Albericio, F. Multifaceted roles of disulfide bonds. Peptides as therapeutics. Chem. Rev. 2014, 114, 901–926. [Google Scholar] [CrossRef]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; McIntosh, J.M. Cloning, synthesis, and characterization of αO-conotoxin GeXIVA, a potent α9α10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci. USA 2015, 112, E4026–E4035. [Google Scholar] [CrossRef] [Green Version]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure–activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Ueberheide, B.M.; Fenyo, D.; Alewood, P.F.; Chait, B.T. Rapid sensitive analysis of cysteine rich peptide venom components. Proc. Natl. Acad. Sci. USA 2009, 106, 6910–6915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougal, O.M.; Turner, M.W.; Ormond, A.J.; Poulter, C.D. Three-dimensional structure of conotoxin tx3a: An M-1 branch peptide of the M-superfamily. Biochemistry 2008, 47, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Dobson, R.; Collodoro, M.; Gilles, N.; Turtoi, A.; Pauw, E.D.; Quinton, L. Secretion and maturation of conotoxins in the venom ducts of Conus textile. Toxicon 2012, 60, 1370–1379. [Google Scholar] [CrossRef]
- Cruz, L.J.; Ramilo, C.A.; Corpuz, G.P.; Olivera, B.M. Conus peptides: Phylogenetic range of biological activity. Biol. Bull. 1992, 183, 159–164. [Google Scholar] [CrossRef]
- Cannon, S.C. Sodium channelopathies of skeletal muscle. Handb. Exp. Pharmacol. 2018, 246, 309–330. [Google Scholar]
- Markgraf, R.; Leipold, E.; Schirmeyer, J.; Paolini-Bertrand, M.; Hartley, O.; Heinemann, S.H. Mechanism and molecular basis for the sodium channel subtype specificity of µ-conopeptide CnIIIC. Br. J. Pharmacol. 2012, 167, 576–586. [Google Scholar] [CrossRef] [Green Version]
- Hameed, S. Nav1. 7 and Nav1. 8: Role in the pathophysiology of pain. Mol. Pain 2019, 15, 1744806919858801. [Google Scholar] [CrossRef] [Green Version]
- Ekberg, J.; Jayamanne, A.; Vaughan, C.W.; Lewis, R.J. μO-conotoxin MrVIB selectively blocks Nav1.8 sensory neuron specific sodium channels and chronic pain behavior without motor deficits. Proc. Natl. Acad. Sci. USA 2006, 103, 17030–17035. [Google Scholar] [CrossRef] [Green Version]
- Breipohl, G.; Knolle, J.; Stuber, W. Synthesis and application of acid labile anchor groups for the synthesis of peptide amides by Fmoc-solid-phase peptide synthesis. Int. J. Pept. Protein Res. 1989, 34, 262–267. [Google Scholar] [CrossRef]
- Heimer, P.; Tietze, A.A.; Bäuml, C.A.; Resemann, A.; Mayer, F.J.; Suckau, D.; Ohlenschläger, O.; Tietze, D.; Imhof, D. Conformational μ-conotoxin PIIIA isomers revisited: Impact of cysteine pairing on disulfide-bond assignment and structure elucidation. Anal. Chem. 2018, 90, 3321–3327. [Google Scholar] [CrossRef]
- Weiser, T. Comparison of the effects of four Na+ channel analgesics on TTX-resistant Na+ currents in rat sensory neurons and recombinant Nav1.2 channels. Neurosci. Lett. 2006, 395, 179–184. [Google Scholar] [CrossRef]
- Mazurek, N.; Byrd, J.C.; Sun, Y.; Ueno, S.; Bresalier, R.S. A galectin-3 sequence polymorphism confers TRAIL sensitivity to human breast cancer cells. Cancer 2011, 117, 4375–4380. [Google Scholar] [CrossRef]
Figure 1.
Sequencing of conotoxin Vi14b by MALDI–TOF–MS/MS analysis.

Figure 2.
MALDI–TOF–MS/MS spectrum of conotoxin Vi002.

Figure 3.
HPLC analysis of stepwise reduction by TCEP and follow-up alkylation by IAM. (A) Stepwise reduction products of Vi14b and Mr3d.1. (B) Alkylated products of stepwise-reduced Vi14b and Mr3d.1.
Figure 3.
HPLC analysis of stepwise reduction by TCEP and follow-up alkylation by IAM. (A) Stepwise reduction products of Vi14b and Mr3d.1. (B) Alkylated products of stepwise-reduced Vi14b and Mr3d.1.

Figure 4.
MALDI–TOF–MS/MS spectrum of product I of Vi14b.

Figure 5.
MALDI–TOF–MS/MS spectrum of product I of Mr3d.1.

Figure 6.
MALDI–TOF–MS/MS spectrum of product II of Mr3d.1.

Figure 7.
Disulfide connectivities of Vi14b, Mr3d.1, Mr3e.1, Mr3.8, Tx3a.1 and TCP.

Figure 8.
Blocking effects of Tx3a.1, Mr3.8, Mr3e.1 and TCP on Nav1.4 and Nav1.8.

Figure 9.
Ovarian cancer cytotoxicity of Tx3a.1, Mr3.8 and Vi14b in ID8 cells (Student’s t-test, * p < 0.05; ** p < 0.01, *** p < 0.001).
Figure 9.
Ovarian cancer cytotoxicity of Tx3a.1, Mr3.8 and Vi14b in ID8 cells (Student’s t-test, * p < 0.05; ** p < 0.01, *** p < 0.001).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Conotoxins identified from Conus venoms.
| Name | Sequence | Origin | Disulfide Bridge | Reference |
|---|---|---|---|---|
| Vi14b | QQMCGRCGGTGQIIK(Ac)NECKTCHGKK(Ac) VTK | C. virgo | 2 pairs | This work |
| Vi002 | LSSGATALSGVPRLTKPAGRLTTTTVAVAF * | C. virgo | none | This work |
| Vi003 | NTESTKGESLLGK * | C. virgo | none | This work |
| Vi15a | DCTTCAGEECCGRCTCPWGDNCSCTEW-(nh2) | C. virgo | 4 pairs | [24] |
| Mr3d.1 | CCRLSCGLGCHPCC | C. marmoreus | 3 pairs | This work |
| Mr3e.1 | VCCPFGGCHELCYCCD | C. marmoreus | 3 pairs | This work |
| Mr3.8 | CCHWNWCDHLCSCCGS | C. marmoreus | 3 pairs | [25,26] |
| Tx3a.1 | CCSWDVCDHPSCTCCG | C. texile | 3 pairs | This work |
| TCP | NCPYCVVYCCPPAYCEASGCRPP | C. texile | 3 pairs | [27] |
* The amino acid “L” in the sequence represented “I/L”, since MS/MS sequencing could not distinguish leucine (L) and isoleucine (I).
Table 2.
Mass of key fragment ions in MS/MS spectrum of conotoxin Vi14b.
| Sequence | ya/yb | Sequence | ya/yb | Sequence | ya/yb |
|---|---|---|---|---|---|
| RCGGT | 475.2082 | TGQIIK(Ac) | 683.4087 | GKK(Ac) | 356.2292 |
| RCGGTG | 504.2347 | IIK(Ac) | 397.2809 | GKK(Ac)V | 427.3027 |
| CGGTGQ | 504.1871 | IK(Ac)N | 398.2398 | GKK(Ac)VT | 528.3504 |
| CGGTGQI | 617.2712 | IK(Ac)NE | 527.2824 | KK(Ac)V | 398.2762 |
| TGQI | 372.2241 | HGK | 323.1826 | K(Ac)VT | 371.2289 |
| IK(Ac) | 284.1969 | GK | 186.1237 |
* yb ion represented the y ion of the intact sequence fragment, while ya was the fragment ion whose C-terminal amino acid residue was decarboxylated.
Table 3.
Mass of key fragment ions in MS/MS spectrum of conotoxin Vi002.
| Sequence | ya/yb | Sequence | ya/yb | Sequence | ya/yb |
|---|---|---|---|---|---|
| SSG | 232.0928 | GVP | 226.1550 | LTT | 316.1867 |
| SSGA | 303.1299 | PR | 226.1662 | AGRLTTT | 673.3992 |
| SSGAT | 404.1776 | LT | 187.1441 | RLTTTT | 674.3832 |
| GATA | 301.1506 | LTK | 315.2391 | TTTV | 403.2187 |
| GATAL | 386.2398 | TKPAG | 455.2613 | TTTVA | 474.2558 |
| GATALS | 473.2718 | PAG | 226.1186 | TTTTVAV | 545.3293 |
| GATALSG | 558.2882 | AGR | 285.1670 | TTVAV | 472.2766 |
| SGVP | 341.1819 | GRL | 299.2190 | VAVA | 341.2183 |
| GV | 129.1022 | LT | 187.1441 |
Table 4.
Current inhibition of the tested conotoxins on Nav1.4 and Nav1.8.
| Name | TP1 | TP2 |
|---|---|---|
| Mr3e.1 | 3.79 ± 0.61% on Nav1.4 | 21.89 ± 3.06% on Nav1.4 |
| Mr3.8 | 4.62 ± 1.06% on Nav1.4 | 23.61 ± 2.70% on Nav1.4 |
| Tx3a.1 | 3.17 ± 0.63% on Nav1.4 | 24.32 ± 2.29% on Nav1.4 |
| TCP | 5.45 ± 0.39% on Nav1.8 | 21.51 ± 1.00% on Nav1.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ju, S.; Zhang, Y.; Guo, X.; Yan, Q.; Liu, S.; Ma, B.; Zhang, M.; Bao, J.; Luo, S.; Fu, Y. Anti-Ovarian Cancer Conotoxins Identified from Conus Venom. Molecules 2022, 27, 6609. https://doi.org/10.3390/molecules27196609
AMA Style
Ju S, Zhang Y, Guo X, Yan Q, Liu S, Ma B, Zhang M, Bao J, Luo S, Fu Y. Anti-Ovarian Cancer Conotoxins Identified from Conus Venom. Molecules. 2022; 27(19):6609. https://doi.org/10.3390/molecules27196609
Chicago/Turabian StyleJu, Shuang, Yu Zhang, Xijun Guo, Qinghui Yan, Siyi Liu, Bokai Ma, Mei Zhang, Jiaolin Bao, Sulan Luo, and Ying Fu. 2022. "Anti-Ovarian Cancer Conotoxins Identified from Conus Venom" Molecules 27, no. 19: 6609. https://doi.org/10.3390/molecules27196609